Bird anatomy, or the physiological structure of birds’ bodies, shows many unique adaptations, mostly aiding flight. Learn about the anatomy of birds
Wing:
FREE Training to Stop Your Bird's Biting
Get FREE access to Beak School – the only system designed to train your bird. Stop behavioral problems like biting, screaming, feather plucking.
Get the Free TrainingMantle: The whole back, combined with the top surface of the wings
Remiges: Large flight feathers on the wings – responsible for supporting the bird during flight. The outer remiges are referred to as the primaries (longest wing feathers). These are the largest, thinnest and stiffest of the flight feathers. The inner remiges – called the secondaries (shorter, upper “arm” feathers) – are attached to the “forearm” (ulna) of a bird.
Rectrices: The long taiI feathers – they help a bird brake and steer in flight. They are in a single horizontal row on the rear margin of the tail.
Coverts or Covert Feathers: Sets of feathers covering other feathers. For example, the wing-coverts are the feathers right above the wing feathers. They cover the base of the flight feathers to provide a smoother surface for the air to flow over.
Undertail Coverts: Feathers around the vent
 Head Anatomy:
Head Anatomy:
 Head Anatomy:
Head Anatomy:- Forehead
- Sinciput
- Septum (= a partition of bone and cartilage between the nasal cavities / nostrils)
- Crown
- Malar Stripe: A line angling back from the bird’s chin, separating the cheek from the throat
- Occiput
- Supercilium
- Eye Stripe / Eye Streak
- Supraloral Line or supraloral feathers of a bird
- Loral area / Lores – The area between beak and eyes
- Cere – an area of soft skin surrounding the nostrils; it may be bare or covered with small, soft feathers
- Epaulets (shoulder patches)
 |
| English: External morphology of a bird (Vanellus malabaricus) BeakHeadIrisPupilMantle (back)Lesser covertsscapulars (shoulder feathers)CovertsTertials (= the flight feathers that are closest to the bird’s body along the wing)RumpPrimaries (longest wing feathers)VentThighTibio-tarsal articulationTarsusFeetTibiaBellyFlanksBreastThroatWattle Français : Morphologie externe d’un oiseau (Vanellus malabaricus) 1 bec2 calotte3 iris4 lore5 manteau6 plumage de petite et moyenne couverture7 plumes scapulaires8 plumage de couverture9 rémiges tertiaires10 croupion11 rémiges primaires12 bas-ventre13 cuisse14 articulation Tibio-tarsale15 tarse16 doigt17 tibia18 ventre19 flanc20 poitrail21 gorge et gorgerin22 caroncule |
Oil Gland is also known as Uropygial Gland or Preen Gland: Located just above the tail.
Bird anatomy, or the physiological structure of birds’ bodies, shows many unique adaptations, mostly aiding flight. Birds have evolved a light skeletal system and light but powerful musculature which, along with circulatory and respiratory systems capable of very high metabolic rates and oxygen supply, permit the bird to fly. The development of a beak has led to evolution of a specially adapted digestive system. These anatomical specializations have earned birds their own class in the vertebrate phylum.
Respiratory system
Due to the high metabolic rate required for flight, birds have a high oxygen demand. Development of an efficient respiratory system enabled the evolution of flight in birds. Birds ventilate their lungs by means of air sacs, structures unique to birds (and hence, perhaps dinosaurs, too). These sacs do not play a direct role in gas exchange, but to store air and act like bellows, allowing the lungs to maintain a fixed volume with fresh air constantly flowing through them.
FREE Training to Stop Your Bird's Biting
Get FREE access to Beak School – the only system designed to train your bird. Stop behavioral problems like biting, screaming, feather plucking.
Get the Free TrainingAir always flows from right (posterior) to left (anterior) through a bird’s lungs during both inhalation and exhalation. Key to a Common Kestrel‘s circulatory lung system: 1 cervical air sac, 2 clavicular air sac, 3 cranial thoracal air sac, 4 caudal thoracal air sac, 5 abdominal air sac (5’ diverticulus into pelvic girdle), 6 lung, 7 trachea
Three distinct sets of organs perform respiration—the anterior air sacs (interclavicular, cervicals, and anterior thoracics), the lungs, and the posterior air sacs (posterior thoracics and abdominals). The posterior and anterior air sacs, typically nine, expand during inhalation. Air enters the bird via the trachea. Half of the inhaled air enters the posterior air sacs, the other half passes through the lungs and into the anterior air sacs. The sacs contract during exhalation. Air from the anterior air sacs empties directly into the trachea and out the bird’s mouth or nares. The posterior air sacs empty their air into the lungs. Air passing through the lungs as the bird exhales is expelled via the trachea. Because fresh air flows through the lungs in only one direction, there is no mixing of oxygen-rich air and oxygen-poor, carbon dioxide-rich, air as in mammalian lungs. Thus, the partial pressure of oxygen in a bird’s lungs is the same as the environment, and so birds have more efficient gas-exchange of both oxygen and carbon dioxide than do mammals.
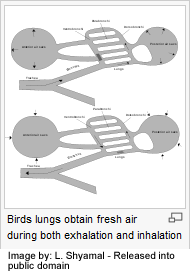
Avian lungs do not have alveoli, as mammalian lungs do, but instead contain millions of tiny passages known as parabronchi, connected at either ends by the dorsobronchi and ventrobronchi. Air flows through the honeycombed walls of the parabronchi into air vesicles, called atria, which project radially from the parabronchi. These atria give rise to air capillaries, where oxygen and carbon dioxide are traded with cross-flowing blood capillaries by diffusion.
Birds also lack a diaphragm. The entire body cavity acts as a bellows to move air through the lungs. The active phase of respiration in birds is exhalation, requiring muscular contraction.
The syrinx is the sound-producing vocal organ of birds, located at the base of a bird’s trachea. As with the mammalian larynx, sound is produced by the vibration of air flowing through the organ. The syrinx enables some species of birds to produce extremely complex vocalizations, even mimicking human speech. In some songbirds, the syrinx can produce more than one sound at a time.

Air always flows from right (posterior) to left (anterior) through a bird’s lungs during both inhalation and exhalation.
Key to a Common Kestrel’s circulatory lung system:
1 cervical air sac
2 clavicular air sac
3 cranial thoracal air sac
4 caudal thoracal air sac
5 abdominal air sac (5′ diverticulus into pelvic girdle)
6 lung
7 trachea
Circulatory System
Birds have four-chambered [heart], in common with humans, most mammals, and some reptiles (namely the crocodilia). This adaptation allows for efficient nutrient and oxygen transport throughout the body, providing birds with energy to fly and maintain high levels of activity. A Ruby-throated Hummingbird‘s heart beats up to 1200 times per minute (about 20 beats per second).
Digestive System
Many birds possess a muscular pouch along the Oesophagus called a crop. The crop functions to both soften food and regulate its flow through the system by storing it temporarily. The size and shape of the crop is quite variable among the birds. Members of the order Columbiformes, such as pigeons, produce a nutritious crop milk which is fed to their young by regurgitation. Birds possess a ventriculus, or gizzard, composed of four muscular bands that rotate and crush food by shifting the food from one area to the next within the gizzard. The gizzard of some species contains small pieces of grit or stone swallowed by the bird to aid in the grinding process of digestion, serving the function of mammalian or reptilian teeth. The use of gizzard stones is a similarity between birds and dinosaurs, which left gizzard stones called gastroliths as trace fossils.
Drinking Behavior
There are four general ways in which birds drink.
Most birds are unable to swallow by the “sucking” or “pumping” action of peristalsis in their esophagus (as humans do), and drink by repeatedly raising their heads after filling their mouths to allow the liquid to flow by gravity, a method usually described as “sipping” or “tipping up”.[4] The notable exception is the Columbiformes; in fact, according to Konrad Lorenz in 1939, “one recognizes the order by the single behavioral characteristic, namely that in drinking the water is pumped up by peristalsis of the esophagus which occurs without exception within the order. The only other group, however, which shows the same behavior, the Pteroclidae, is placed near the doves just by this doubtlessly very old characteristic.”
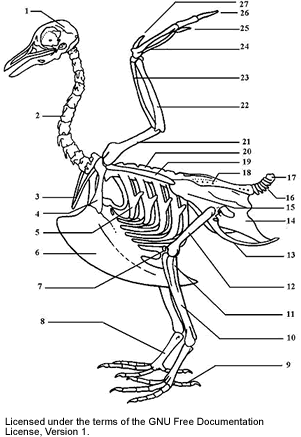 |
| A stylised bird skeleton skull cervical vertebrae furcula coracoid uncinate process keel patella tarsometatarsus digits tibiotarsus (long leg bone) (10 and 11) tibiotarsus (10 and 11) femur ischium (innominate bone) pubi (innominate bone) illium (innominate bone) caudal vertebrae pygostyle synsacrum scapula lumbar vertebrae humerus ulna radius carpus metacarpus digits alula |
Although this general rule still stands, since that time, observations have been made of a few exceptions in both directions.
FREE Training to Stop Your Bird's Biting
Get FREE access to Beak School – the only system designed to train your bird. Stop behavioral problems like biting, screaming, feather plucking.
Get the Free TrainingIn addition, specialized nectar feeders like sunbirds (Nectariniidae) and hummingbirds (Trochilidae) drink by using protrusible grooved or trough-like tongues, and parrots (Psittacidae) lap up water.
Skeletal System
The bird skeleton is highly adapted for flight. It is extremely lightweight but strong enough to withstand the stresses of taking off, flying, and landing. One key adaptation is the fusing of bones into single ossifications, such as the pygostyle. Because of this, birds usually have a smaller number of bones than other terrestrial vertebrates. Birds also lack teeth or even a true jaw, instead having evolved a beak, which is far more lightweight. The beaks of many baby birds have a projection called an egg tooth, which facilitates their exit from the amniotic egg.
Birds have many bones that are hollow with criss-crossing struts or trusses for structural strength. The number of hollow bones varies among species, though large gliding and soaring birds tend to have the most. Respiratory air sacs often form air pockets within the semi-hollow bones of the bird’s skeleton. Some flightless birds like penguins and ostriches have only solid bones, further evidencing the link between flight and the adaptation of hollow bones.
Birds also have more cervical (neck) vertebrae than many other animals; most have a highly flexible neck consisting of 13-25 vertebrae. Birds are the only vertebrate animals to have a fused collarbone (the furcula or wishbone) or a keeled sternum or breastbone. The keel of the sternum serves as an attachment site for the muscles used for flight, or similarly for swimming in penguins. Again, flightless birds, such as ostriches, which do not have highly developed pectoral muscles, lack a pronounced keel on the sternum. It is noted that swimming birds have a wide sternum, while walking birds had a long or high sternum while flying birds have the width and height nearly equal.
Birds have uncinate processes on the ribs. These are hooked extensions of bone which help to strengthen the rib cage by overlapping with the rib behind them. This feature is also found in the tuatara Sphenodon. They also have a greatly elongate tetradiate pelvis as in some reptiles. The hindlimb has an intra-tarsal joint found also in some reptiles. There is extensive fusion of the trunk vertebrea as well as fusion with the pectoral girdle. They have a diapsid skull as in reptiles with a pre-lachrymal fossa (present in some reptiles). The skull has a single occipital condyle.
Skeletal composition
The skull consists of five major bones: the frontal (top of head), parietal (back of head), premaxillary and nasal (top beak), and the mandible (bottom beak). The skull of a normal bird usually weighs about 1% of the birds total bodyweight.
 The vertebral column consists of vertebrae, and is divided into three sections: cervical (13-16) (neck), Synsacrum (fused vertebrae of the back, also fused to the hips (pelvis), and pygostyle (tail).
The vertebral column consists of vertebrae, and is divided into three sections: cervical (13-16) (neck), Synsacrum (fused vertebrae of the back, also fused to the hips (pelvis), and pygostyle (tail).
The chest consists of the furcula (wishbone) and coracoid (collar bone), which two bones, together with the scapula (see below), form the pectoral girdle. The side of the chest is formed by the ribs, which meet at the sternum (mid-line of the chest).
The shoulder consists of the scapula (shoulder blade), coracoid (see The Chest), and humerus (upper arm). The humerus joins the radius and ulna (forearm) to form the elbow. The carpus and metacarpus form the “wrist” and “hand” of the bird, and the digits (fingers) are fused together. The bones in the wing are extremely light so that the bird can fly more easily.
The hips consist of the pelvis which includes three major bones: Illium (top of the hip), Ischium (sides of hip), and Pubis (front of the hip). These are fused into one (the innominate bone). Innominate bones are evolutionary significant in that they allow birds to lay eggs. They meet at the acetabulum (the hip socket) and articulate with the femur, which is the first bone of the hind limb.
The upper leg consists of the femur. At the knee joint, the femur connects to the tibiotarsus (long leg bone – shin) and fibula (side of lower leg). The tarsometatarsus forms the upper part of the foot, digits make up the toes. The leg bones of birds are the heaviest, contributing to a low center of gravity. This aids in flight. A bird’s skeleton comprises only about 5% of its total body weight
Birds feet are classificated as anisodactyl, zygodactyl, heterodactyl, syndactyl or pamprodactyl.[9]
Muscular system
Most birds have approximately 175 different muscles, mainly controlling the wings, skin, and legs. The largest muscles in the bird are the pectorals, or the breast muscles, which control the wings and make up about 15 – 25% of a flighted bird’s body weight. They provide the powerful wing stroke essential for flight. The muscle ventral (underneath) to the pectorals is the supracoracoideus. It raises the wing between wingbeats. The supracoracoideus and the pectorals together make up about 25 – 35% of the bird’s full body weight.
The skin muscles help a bird in its flight by adjusting the feathers, which are attached to the skin muscle and help the bird in its flight maneuvers.
There are only a few muscles in the trunk and the tail, but they are very strong and are essential for the bird. The pygostyle controls all the movement in the tail and controls the feathers in the tail. This gives the tail a larger surface area which helps keep the bird in the air.

Head
Birds have acute eyesight – raptors have vision eight times sharper than humans – thanks to higher densities of photoreceptors in the retina (up to 1,000,000 per square mm in Buteos, compared to 200,000 for humans), a high number of optic nerves, a second set of eye muscles not found in other animals, and, in some cases, an indented fovea which magnifies the central part of the visual field. Many species, including hummingbirds and albatrosses, have two foveas in each eye. Many birds can detect polarised light. The eye occupies a considerable part of the skull and is surrounded by a sclerotic eye-ring, a ring of tiny bones that surround the eye. This character is also seen in the reptiles.
The bills of many waders have Herbst corpuscles which help them detect prey hidden under wet sand using minute pressure differences in the water. All extant birds can move the parts of the upper jaw relative to the brain case. However this is more prominent in some birds and can be readily detected in parrots.[11]
Birds have a large brain to body mass ratio. This is reflected in the advanced and complex bird intelligence.
The region between the eye and bill on the side of a bird’s head is called the lore. This region is sometimes featherless, and the skin may be tinted, as in many species of the cormorant family.
Reproduction
Although most male birds have no external sex organs, the male does have two testes which become hundreds of times larger during the breeding season to produce sperm.[12] The female’s ovaries also become larger, although only the left ovary usually functions. However, if the left ovary is damaged by infection or other problems, the right ovary will try to function.
In the males of species without a phallus (see below), sperm is stored in the seminal glomera within the cloacal protuberance prior to copulation. During copulation, the female moves her tail to the side and the male either mounts the female from behind or in front (in the stitchbird), or moves very close to her. The cloacae then touch, so that the sperm can enter the female’s reproductive tract. This can happen very fast, sometimes in less than half a second.
The sperm is stored in the female’s sperm storage tubules for a week to a year, depending on the species. Then, eggs will be fertilised individually as they leave the ovaries, before being laid by the female. The eggs continue their development outside the female body.
Many waterfowl and some other birds, such as the ostrich and turkey, possess a phallus. When not copulating, it is hidden within the proctodeum compartment within the cloaca, just inside the vent.
After the eggs hatch, parents provide varying degrees of care in terms of food and protection. Precocial birds can care for themselves independently within minutes of hatching; altricial hatchlings are helpless, blind, and naked, and require extended parental care. The chicks of many ground-nesting birds such as partridges and waders are often able to run virtually immediately after hatching; such birds are referred to as nidifugous. The young of hole-nesters, on the other hand, are often totally incapable of unassisted survival. The process whereby a chick acquires feathers until it can fly is called “fledging”.
Some birds, such as pigeons, geese, and Red-crowned Cranes, remain with their mates for life and may produce offspring on a regular basis.
Scales
The scales of birds are composed of the same keratin as beaks, claws, and spurs. They are found mainly on the toes and metatarsus, but may be found further up on the ankle in some birds. Most bird scales do not overlap significantly, except in the cases of kingfishers and woodpeckers. The scales of birds are thought to be homologous to those of reptiles and mammals.
Bird embryos begin development with smooth skin. On the feet, the corneum, or outermost layer, of this skin may keratinize, thicken and form scales. These scales can be organized into;
- Cancella – minute scales which are really just a thickening and hardening of the skin, crisscrossed with shallow grooves.
- Reticula – small but distinct, separate, scales. Found on the lateral and medial surfaces (sides) of the chicken metatarsus. These are made up of alpha-keratin.
- Scutella – scales that are not quite as large as scutes, such as those found on the caudal, or hind part, of the chicken metatarsus.
- Scutes – the largest scales, usually on the anterior surface of the metatarsus and dorsal surface of the toes. These are made up of beta-keratin as in reptilian scales.
The rows of scutes on the anterior of the metatarsus can be called an acrometatarsium or acrotarsium.
Feathers can be intermixed with scales on some birds’ feet. Feather follicles can lie between scales or even directly beneath them, in the deeper dermis layer of the skin. In this last case, feathers may emerge directly through scales, and be encircled at the plane of emergence entirely by the keratin of the scale.
Bird Anatomy Glossary
Sinciput
Sinciput is the upperpart of the skull – it includes the forehead and the crown
Occiput
Together with the quadrates, the opisthotics and various dermal bones, the bones of the occiput form the back of a bird’s skull.
Supercilium
Supercilium – also commonly referred to as “eyebrow” — is a stripe which starts above the bird’s loral area (area between beak and eyes), continuing above the eye, and finishing somewhere towards the rear of the bird’s head.
FREE Training to Stop Your Bird's Biting
Get FREE access to Beak School – the only system designed to train your bird. Stop behavioral problems like biting, screaming, feather plucking.
Get the Free TrainingWhere a stripe is present only above the lores, and does not continue behind the eye, it is called a supraloral stripe or simply supraloral.
The Supercilium is distinct from the eyestripe which is a line which runs across the lores, and continues behind the eye.
Copyright: Wikipedia. This article is licensed under the GNU Free Documentation License. It uses material from the Wikipedia.org.
Index of Bird Diseases .. Symptoms and Potential Causes … Bird Species and Diseases They are Most Susceptible to … Bird Health Care
NEED A VET?
USA: Find Your Local Avian Veterinarian
Information contained on this website is provided as general reference only. For application to specific circumstances, professional advice should be sought.
Please Note: The articles or images on this page are the sole property of the authors or photographers. Please contact them directly with respect to any copyright or licensing questions. Thank you.